

これまでに提唱されたいろいろな血液細胞分化経路モデル
2009年2月4日
河本 宏
血液学や免疫学の教科書には、30年も前から大同小異の造血モデルが描かれていた。それは、最初の分岐でリンパ球系列と赤血球/ミエロイド系列に分かれるという図である。一方、われわれはこれまでに独自の「ミエロイド基本型モデル」を提唱してきた。これら2つのモデル以外にも、歴史的には、いろいろな造血分化経路モデルが提唱されて来た。本稿では、それらのモデルを紹介する。
まず、語句の整理をしておこう。ミエロイド系は、単球/マクロファージと顆粒球を含む。なお、この2種類の細胞は常にセットになっているとは限らないことに注意していただきたい。すなわち、ある前駆細胞がマクロファージしかつくらない場合でも「ミエロイド系細胞への分化能を有する」と表す。巨核球系細胞は、本稿では簡略化のためエリスロイド系列に含めて扱う。すなわち、血液細胞を大きくエリスロイド系(E)、ミエロイド系(M)、T細胞系(T)、B細胞系(B)に分けて考える。
では、これまでに提唱されて来た主な血液細胞分化経路モデルを、提唱された順にみていこう。
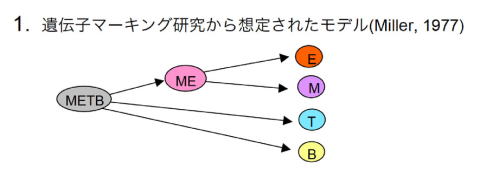
1977年に、放射線照射で誘発した染色体異常をマーカーとして観察する方法で、T、B、ミエロイド系細胞の3系列が同じ細胞由来であることが示された[1]。この研究では、上記3系列のうち2つの系列が共通してマークされるというデータは得られなかったので、多能幹細胞からT、B、ミエロイド系細胞への分化経路は、それぞれ独立に起こると考えられた。ME共通細胞が描かれているが、これには根拠があった訳ではない。当時は、ミエロイド系幹細胞は存在するものと思われていたので、こういう描き方になったようである。
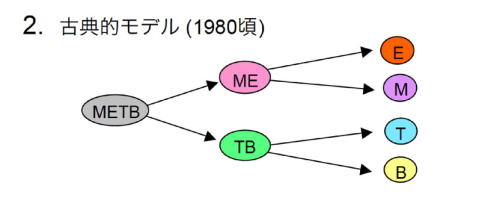
上記の実験で研究でT-B共通の前駆細胞がみつかってなかったにもかかわらず、80年代以後の総説や教科書では、古典的モデルがよく使われるようになった。最初の分岐でME前駆細胞(common myelo-eruthroid progenitor; CMLP)とTB前駆細胞(common lymphoid progenitor, CLP)がつくられるとしている。実証されてもいないのにこのモデルが使われてきたのは、T細胞とB細胞が形態的に似ているということと、いずれもが抗原特異的な免疫反応のプレーヤーとして働くという、他の細胞とは決定的に異なる特徴を有しているゆえに、「T細胞とB細胞は近縁である」と考えられたからである。
1997年にI.L. Weissmanのグループはマウス骨髄のLineageマーカー陰性ckit弱陽性IL-7R陽性分画にCLPをみつけたと報告した [2]。これは、古典的モデルを支持するものだった。しかし、このIL-7R陽性細胞はin vitroではミエロイド細胞をよく生成するし [3,4]、また生体中でミエロイド細胞を生成しないということが厳密に証明されている訳ではない。また、生理的にはB細胞の前駆細胞として働いていて、胸腺に移行してT細胞をつくることはないと考えられている[5,6]。
一方、CMLPについても、2000年にWeissmanのグループが検出したと報告した [7]。ただし、CMLPがミエロイド系細胞をつくるための主な経路であるかどうかは示されていない。
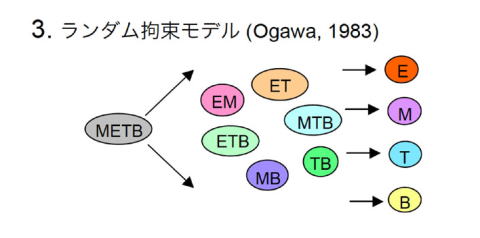
M. Ogawaらによって1983年に提唱されたモデル [8]。確率的モデルとも呼ばれている。コロニー法で骨髄細胞を解析するといろいろな組み合わせで複数種の細胞から成るコロニーが形成されるという知見に準拠したもので、「系列決定はランダムな組み合わせで進行する」というコンセプトを提示している。元来はミエロ-エリスロイド系の範囲内で提唱されたモデルであったが、敷衍してT/B系列も含めて解釈する研究者も多くいた。発生生物学の原理からかなりはずれた奇妙なモデルであるが、当時はおもしろいモデルとして一定の評価を得ていたようで、特に本邦では一世を風靡した。その後の研究で組み合わせには一定のルールがあることが明らかになり、今日ではこのモデルの意義は失われている。
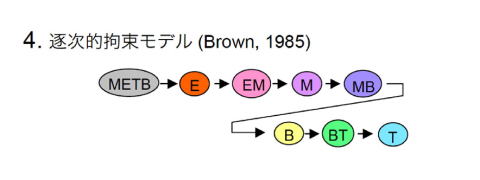
G. Brownらが1985年に提唱したモデルでは、多能前駆細胞は、分化に従ってエリスロイド、ミエロイド、B、Tというように順次分化能が変わり、その途中、中間的な前駆細胞もつくられるとされた [9]。B細胞とミエロイド細胞が近縁であることは当時から指摘されていたので、その知見を組み込もうとしたモデルである。おもしろいモデルだが、例えば後述するミエロイド-T前駆細胞の存在はこのモデルでは説明できない。
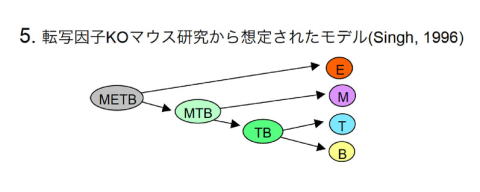
1996年に転写因子欠損マウスの解析からH. Singhが提唱したモデル [10]。このモデルの主な論拠となっているのは、Pu.1欠損マウスで赤血球/血小板は正常なのにミエロイド、T、B系列が欠如するとした報告と、Ikaros欠損マウスでT、B系列だけが欠如するとした報告である [11,12]。しかし、これらの表現型解析結果は、その後の詳しい研究により不正確であったことがわかっている [13,14]。Pu.1欠損マウス研究から得られたミエロイド系とリンパ系の結びつきが深いという洞察自体は正しかったと思われるが、Ikaros欠損マウスについては、古典的モデルに合わせて無理な解釈がなされたように見受けられる。
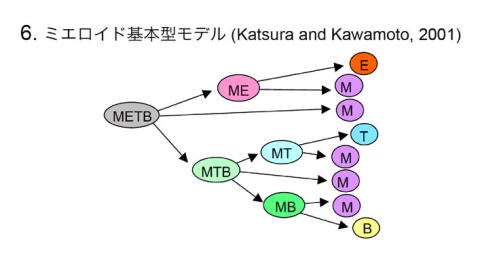
われわれは1997年頃からクローナルアッセイを用いて系列決定過程を明らかにしてきた [15-18]。それらのデータを総合して、2001年に新しいモデルとして提唱した [19-22]。このモデルでは、最初の分岐点でCMEPとCMLPが生成する(図1b)。CMLPから、ミエロイド-T前駆細胞(MTP)およびミエロイド-B前駆細胞(MBP)という分化段階を経て、TおよびB前駆細胞がつくられる。古典的モデルでは全ての系列を同等に扱っているが、ミエロイド基本型モデルでは、ミエロイド系列を基本型、他の系列を特殊化した系列ととらえている点で、本質的に異なる。このモデルは胎生期造血を表しているという意見も出されていた [23,24]が、最近のわれわれが報告したT前駆細胞がミエロイド系細胞をつくるという知見 [25]は、成体造血においてもこのモデルを強く支持するものであった。
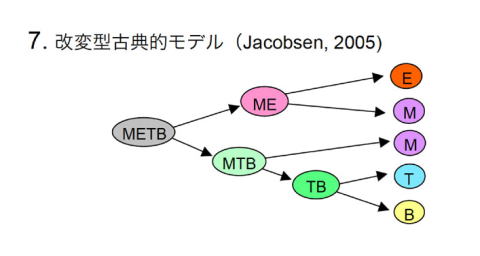
2005年に、S. Jacobsonらによって骨髄中にCMLP(MTB前駆細胞)を同定したという報告がなされた [26]。2002年に筆者らが胎仔肝臓で同定したCMLP [18]の、骨髄におけるカウンターパートにあたる。この知見は筆者らのミエロイド基本型モデルに合致しているのだが、Jacobsonらの提唱するモデルではCMLPを造血幹細胞(METB)とCLP(TB)の間に位置する細胞とした。すなわち、コンセプト的には古典的モデルの域にとどまっていた。
文献
1 Abramson, S. et al. (1977) The identification in adult bone marrow of pluripotent and restricted stem cells of the myeloid and lymphoid systems. J. Exp. Med 145, 1567-1569
2 Kondo, M. et al. (1997) Identification of clonogenic common lymphoid progenitors in mouse bone marrow. Cell 91, 661-672
3 Balciunaite, G. et al. (2005) A B220+ CD117+ CD19- hematopoietic progenitor with potent lymphoid and myeloid developmental potential. Eur. J. Immunol. 35, 2019-2030
4 Rumfelt, L.L. et al. (2006) Lineage specification and plasticity in CD19- early B cell precursors. J. Exp. Med. 203, 675-687
5 Allman, D. et al. (2003) Thymopoiesis independent of common lymphoid progenitors. Nat. Immunol. 4, 168-174
6 Borghesi, L. et al. (2004) B lineage-specific regulation of V(D)J recombinase activity is established in common lymphoid progenitors. J. Exp. Med. 199, 491-502
7 Akashi, K. et al. (2000) A clonogenic common myeloid progenitor that gives rise to all myeloid lineages. Nature 404, 193-197
8 Ogawa, M. et al. (1983) Renewal and commitment to differentiation of hemopoietic stem cells (an interpretive review). Blood 61, 823-829
9 Brown, G. et al. (1985) Sequential determination of lineage potentials during haemopoiesis. Br. J. Cancer 52, 681-686
10 Singh, H. (1996) Gene targeting reveals a hierarchy of transcription factors regulating specification of lymphoid cell fates. Curr. Opin. Immunol. 8, 160-165
11 Scott, E.W. et al. (1994) Requirement of transcription factor PU.1 in the development of multiple hematopoietic lineages. Science 265, 1573-1577
12 Georgopoulos, K. et al. (1994) The Ikaros gene is required for the development of all lymphoid lineages. Cell 79, 143-156
13 Wang, J.H. et al. (1996) Selective defects in the development of the fetal and adult lymphoid system in mice with an Ikaros null mutation. Immunity 5, 537-549
14 Spain, L.M. et al. (1999) T cell development in PU.1-deficient mice. J. Immunol. 163, 2681-2687
15 Kawamoto, H. et al. (1997) Direct evidence for the commitment of hematopoietic stem cells to T, B and myeloid lineages in murine fetal liver. Int. Immunol. 9, 1011-1019
16 Kawamoto, H. et al. (1999) Emergence of T cell progenitors without B cell or myeloid differentiation potential at the earliest stage of hematopoiesis in the murine fetal liver. J. Immunol. 162, 2725-2731
17 Kawamoto, H. et al. (2000) T cell progenitors emerge earlier than B cell progenitors in the murine fetal liver. Immunity 12, 441-450
18 Lu, M. et al. (2002) The common myelolymphoid progenitor: a key intermediate stage in hemopoiesis generating T and B cells. J. Immunol. 169, 3519-3525
19 Katsura, Y. and Kawamoto, H. (2001) Stepwise lineage restriction of progenitors in lympho-myelopoiesis. Int. Rev. Immunol. 20, 1-20
20 Katsura, Y. (2002) Redefinition of lymphoid progenitors. Nat. Rev. Immunol. 2, 127-132
21 Kawamoto, H. (2006) A close developmental relationship between the lymphoid and myeloid lineages. Trends Immunol. 27, 169-175
22 Kawamoto, H, and Katsura, Y. New paradigm for hematopoietic cell lineages: revision of the classical concept of myeloid-lymphoid dichotomy. Trends Immunol. in press.
23 Laiosa, C.V. et al. (2006) Determinants of lymphoid-myeloid lineage diversification. Annu. Rev. Immunol. 24, 705-738
24 Buza-Vidas, N. et al. (2007) Delineation of the earliest lineage commitment steps of haematopoietic stem cells: new developments, controversies and major challenges. Curr. Opin. Hematol. 14, 315-321
25 Wada, H. et al. (2008) Adult T-cell progenitors retain myeloid potential. Nature 452, 768-772
26 Adolfsson, J. et al. (2005) Identification of Flt3+ lympho-myeloid stem cells lacking erythro-megakaryocytic potential a revised road map for adult blood lineage commitment. Cell 121, 295-306